
Efforts to understand how intrinsically disordered regions interact have produced a variety of answers

It’s a bit ironic that, just as methods for deducing the crystal structures of protein molecules have become supercharged, we’ve started to discover they might not always help very much for developing drugs that target proteins. First there was synchrotron radiation, and now free-electron lasers, as sources of ultrabright x-ray beams that can reduce data collection times and crystal sizes for crystallography.1 Meanwhile, AlphaFold can predict the structures of many proteins with impressive accuracy from sequence data alone.
Yet it’s not always a question of structure. Take a recent study by a team at University College London, UK, revealing the crucial role of dynamics and of the ability of enzymes to explore their conformational landscape.2 The researchers point out that some single-residue mutations of proteins can significantly change their activity while barely impacting their crystal structure, because the active conformations are only sparsely represented in the distribution of dynamical states.
Intrinsically disordered regions
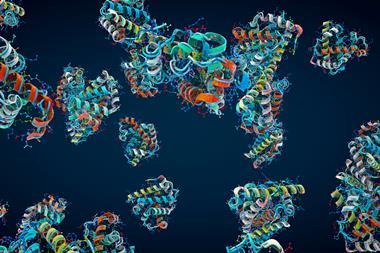
A still more fundamental limitation to the ability of structure to determine function (and thus to suggest which drug molecules might interfere with it) arises when there is no structure to speak of. Many proteins, including some of those most central to cell behaviour – such as transcription factors that govern gene regulation – have considerable amounts of structural disorder. They are said to contain intrinsically disordered regions, which sometimes constitute almost the entire protein.3 These molecules tend to be floppy in solution, lacking the characteristic secondary structures such as alpha helices or beta sheets, and are hard to crystallise.
Their protean nature typically makes proteins like this rather unselective about which other molecules they bind. This probably explains why proteins that sit at some hubs of the cell’s interaction networks – like the tumour suppressor p53, which regulates transcription and is implicated in around half of all human cancers – have a high degree of disorder. Such flexibility permits the protein to forge new connections in the network, potentially contributing to the evolvability of the parent organism.
Because intrinsically disordered regions make proteins apt to associate and aggregate rather indiscriminately, it’s no surprise either that such proteins feature in the formation both of condensates – dense blobs of proteins that phase-separate within cells and seem crucial for a range of functions from gene regulation to RNA splicing4 – and of cytotoxic aggregates linked to neurodegenerative diseases such as Alzheimer’s. For this reason, intrinsically disordered proteins could be important targets for drugs.
A team of researchers primarily based in Cambridge, UK, has now developed a computational scheme for identifying which proteins of this sort might be best targeted to intervene in condensate formation.5 They’ve tested it out on three such proteins, looking at how such interventions affect the aggregation behaviour in model cell systems.
Methods like this, assisted by machine learning, might identify candidate drug targets by crunching the data. But as with drug development more generally, researchers are likely to want some understanding of the principles behind these molecular interactions, to introduce an element of rational design.
That’s hard, because intrinsically disordered proteins lack the ‘lock-and-key’ binding selectivity seen for ordered proteins – although some adopt a more ordered structure when they bind.6 So if shape isn’t the determining factor, what is? One suggestion is that proteins involved in condensate formation use ‘sticky patches’ along their chains to form many weak associations.7 Now several recent papers have taken a closer look at particular interactions between intrinsically disordered regions to get at the general thermodynamic principles involved.
In the balance
As with any biomolecular interactions, there’s a balance between enthalpic contributions – alignment of groups that attract one another, say – and entropic ones due to the expulsion and ‘liberation’ of solvation water or the decreased mobility of bound chains. But at face value the new studies seem to arrive at different conclusions about what matters most. Debasis Saha and Biman Jana of the Indian Association for the Cultivation of Science in Kolkata investigated the factors governing the dimerisation of like-charged model peptides (assuming this crudely approximates the aggregation within condensates) and conclude that positively charged peptides are best for making the dense phase.8
Meanwhile, Benjamin Schuler at the University of Zurich, Switzerland, and colleagues looked at the interaction between a pair of positively and negatively charged disordered proteins known to interact in vivo and see a large entropic contribution to binding coming from the release of counterions.9 And Saumyak Mukherjee and Lars Schäfer at the Ruhr University Bochum, Germany, concluded that the entropic effects of solvation-water release from the dense condensate are at least as important as protein–protein enthalpic interactions.10
What to conclude from all this? Well, it wouldn’t be surprising if disordered proteins mobilise a range of different interactions to do their jobs: the devil is in the details. The big question is whether those details can be harnessed to control these collective and fuzzy interactions – and cure disease.
References
1. I Poudyal, M Schmidt and P Schwander, Struct. Dyn., 2020, 7, 024102 (DOI: 10.1063/1.5144516)
2. V K Shukla, L. Siemons and D F Hansen, PNAS, 2023 120, e2310910120 (DOI: 10.1073/pnas.2310910120)
3. H J Dyson and P E Wright, Nat. Rev. Mol. Cell Biol., 2005, 6, 197 (DOI: 10.1038/nrm1589)
4. Y Shin and C P Brangwynne, Science, 2017, 357, eaaf4382 (DOI: 10.1126/science.aaf4382)
5. C M Lim et al., PNAS, 2023, 120, e2300215120 (DOI: 10.1073/pnas.2300215120)
6. V N Uversky, Adv. Protein Chem. Struct. Biol., 2018, 110, 85 (DOI: 10.1016/bs.apcsb.2017.06.005)
7. K M Ruff et al., Mol. Cell, 2022, 82, 3198 (DOI: 10.1016/j.molcel.2022.06.024)
8. D Saha and B Jana, J. Phys. Chem. B, 2023, 127, 6656 (DOI: 10.1021/acs.jpcb.3c03087)
9. A Chowdhury et al., PNAS, 2023, 120, e2304036120 (DOI: 10.1073/pnas.2304036120)
10. S Mukherjee and L Schäfer, Nat. Commun., 2023, 14, 5892 (DOI: 10.1038/s41467-023-41586-y)











No comments yet