
The discovery of the importance and structure DNA was more than just Crick and Watson’s eureka moment. Mike Sutton untangles the tale of life’s molecular mysteries
The discovery of the importance and structure DNA was more than just Crick and Watson’s eureka moment. Mike Sutton untangles the tale of life’s molecular mysteries

When their search began, the composition of nucleic acids was already known. In 1869 the Swiss biologist Friedrich Miescher extracted an acidic substance rich in phosphorus from the nuclei of blood cells, and called it ‘nuclein’. It contained protein, and another substance later described as ‘nucleic acid’. Different sources yielded variants, including ‘yeast nucleic acid’, subsequently identified as ribonucleic acid (RNA) and ‘pancreas nucleic acid’, now known to be deoxyribonucleic acid (DNA).
Between 1895 and 1901 the German biochemist Albrecht Kossel isolated five organic bases – aadenine, guanine, cytosine thymine and uracil – from nucleic acids. The picture was further clarified by Phoebus Levene, the head of biochemistry at New York’s Rockefeller Institute from 1905. He showed that the huge nucleic acid molecules could be broken into much smaller units which he called ‘nucleotides’.
Every nucleotide has three components – a sugar molecule, an acidic phosphate group and one of Kossel’s five nucleobases. In RNA, the sugar is ribose, and the four possible bases are adenine, guanine, cytosine, and uracil. DNA’s sugar is deoxyribose while its bases are adenine, guanine, cytosine and thymine. In both, alternate sugar molecules and phosphates form a chain connected by ester linkages.
Genetic suspect
Levene estimated that the nucleobases occurred in approximately equal numbers. This prompted the (incorrect) assumption that DNA consisted of many repetitions of a four-nucleotide unit containing one molecule of each base. Biologists considered this structure too simple to carry the genetic information passed on when cells divide, and assumed the nucleoproteins performed that function. Nucleic acids were thought to play an auxiliary role – perhaps as scaffolding to support fragile protein molecules.
By the late 1940s, however, a few researchers suspected that the instructions which made an acorn grow into an oak tree – rather than an elm or an elephant – were written in nucleic acid molecules. Several of these investigators were inspired by What is Life? – a speculative book published in 1944 by the Austrian physicist Erwin Schrödinger. It focussed on the fuzzy frontier between very small living organisms and very large non-living molecules. However, the unification of these two realms seemed unachievable until fresh clues emerged from previously ignored or undervalued laboratory results.The conclusion that DNA was the genetic information carrier had profound implications
Fred Griffith was a pathologist working for Britain’s Ministry of Health. In 1928 he mixed dead bacteria from a virulent pneumonia strain with live ones of a similar but non-virulent type, and injected mice with the mixture. They became fatally infected with live bacteria of the virulent strain. Griffith argued that an unknown chemical substance – the ‘transforming principle’ – had passed genetic information from dead bacteria to living ones, enabling them to breed new copies of the lethal variant.
Other bacteriologists – including Oswald Avery at the Rockefeller Institute – were sceptical, because cross-contamination was a familiar hazard in this type of research. However, Avery’s team eventually confirmed Griffith’s results, and accomplished similar bacterial transformations without using animal hosts. After many experiments, Avery and his associates suggested in 1944 that Griffith’s transforming principle was DNA. At first their paper attracted little interest – partly because by then much of the scientific community was involved in war work.
X-rays at King’s
British scientists recruited for military research included Francis Crick, who helped the Royal Navy improve its anti-mine defences, and Maurice Wilkins, who worked on uranium isotope separation for the Manhattan project. Both returned to academia disillusioned with physics, but inspired by Schrödinger’s book. Crick joined the Medical Research Council’s (MRC) molecular biology unit at Cambridge, while Wilkins settled in the biophysics unit at King’s College London.

In May 1950 King’s received a high-quality sample of DNA’s sodium salt from the Swiss organic chemist Rudolf Signer. Wilkins used a portion for optical analysis, and passed some on to Raymond Gosling, a PhD student working on X-ray diffraction. Gosling’s early X-ray photographs were disappointing, but Wilkins had discovered how to make long DNA fibres, from which he and Gosling eventually obtained much clearer X-ray diffraction patterns.
Their pictures confirmed that DNA fibres which had absorbed an appropriate amount of water exhibited a relatively orderly structure – a deduction already made from poorer photographs by William Astbury at Leeds University. Viewed alongside Avery’s (still controversial) conclusion that DNA was the genetic information carrier, this result had profound implications. Naturally, Wilkins hoped better X-ray pictures would reveal more about DNA’s structure. So did his friend Francis Crick.
In January 1951, King’s College acquired a new research fellow. Rosalind Franklin was a Cambridge-educated chemist who had spent the war employed by the Coal Utilisation Research Association. Later she worked at a government research laboratory in Paris, acquiring considerable expertise in X-ray diffraction there. The head of biophysics at King’s, John Randall, immediately assigned the X-ray work to her, along with Gosling’s PhD supervision and Signer’s DNA sample.
Wilkins was not consulted about the change, and the inevitable uneasiness between him and Franklin was amplified by differences in temperament. Nevertheless, Wilkins continued working on DNA acquired from another source. At a conference in Naples in May 1951 his results aroused little interest. But his audience included an American research student who believed this investigation might start a revolution. Wilkins’ cool response to his youthful enthusiasm did not stop him wanting to join it.
An American in Cambridge
James Watson had entered the University of Chicago at 15, and left Indiana University with a PhD at 22. Along the way his focus moved from ornithology to bacteriology, and he encountered Schrödinger’s What is Life? His supervisor, Salvador Luria, found him a post-doctoral project in Copenhagen where he did useful, though unexciting, lab work on bacterial genetics until the Naples symposium steered him in a new direction.
On returning from Naples to Denmark, Watson learned that America’s most celebrated chemist, Linus Pauling, had announced a dramatic discovery. Applying his unrivalled knowledge of bond lengths and bond angles, Pauling showed how a long polypeptide molecule could twist into a spiral, stabilised by hydrogen bonds linking one coil to the next. His model – the alpha-helix – appeared to be compatible with existing crystallographic evidence about protein structures.
Watson hoped to achieve a similar result with DNA – but first he needed to learn more chemistry (he had been excluded from the lab at Indiana University after trying to heat benzene with a Bunsen burner!). In October 1951, with Luria’s endorsement, he joined the MRC group led by John Kendrew at Cambridge, studying the muscle protein myoglobin. There he encountered Crick, who was pursuing a PhD on protein crystallography under Max Perutz. After discovering their shared interest in DNA, Crick and Watson began investigating it unofficially.
Mounting evidence
Meanwhile at King’s, Franklin and Gosling gradually improved their apparatus and techniques. They confirmed that the long fibres of ‘wet’ DNA (known as the B-form) gave X-ray diffraction patterns different from the ‘dry’ or A-form, indicating that hydration altered the molecule’s shape. Although Franklin recognised that a helical structure for DNA was possible, she regarded speculation as premature while evidence remained so limited. Spurning Pauling’s model-building approach, she preferred to expose the DNA molecule’s geometry slowly but surely, through the mathematical analysis of X-ray diffraction patterns.

In November 1951 Franklin led a seminar on the DNA work at King’s, which Watson attended. Having taken no notes, he left with the erroneous impression that Franklin entirely rejected a helical structure for DNA. Moreover, to Crick’s annoyance, he forgot her figures for the B-form’s water content. But despite their limited data, the Cambridge duo continued model-building, worried that Pauling might find the structure first.
Just before Christmas 1951 they produced a structure that seemed satisfactory. It had three DNA chains twisted into a spiral, with their sugar–phosphate backbones inside and the nucleobases pointing outwards. Triumphantly, they invited Wilkins to view it. He did – but Franklin came with him, and quickly demonstrated that their structure was chemically unfeasible
Crick and Watson thought the intertwined DNA strands were knitted together by weak bonds between phosphate groups and adjacent metallic cations. This form of linkage – known as ‘salt bridging’ – seemed plausible only because of Watson’s inaccurate recollection of the B-form’s water content. The actual degree of hydration was higher than he supposed, meaning that there would be far too many water molecules around the ions for salt bridges to form.
Franklin regarded this debacle as a vindication of her cautious approach to the puzzle, though Wilkins was less certain. Meanwhile Crick and Watson were told by their superiors to drop the topic. And so in January 1952 Crick returned to his protein studies, while Watson took X-ray diffraction pictures of tobacco mosaic virus. Nevertheless, DNA was seldom far from their thoughts.
The race gathers pace
In the spring of 1952 they learned of a significant breakthrough, made at New York’s Cold Spring Harbour Laboratory by Alfred Hershey and Martha Chase. After introducing a radioactive phosphorus isotope into the DNA of a bacteriophage (a virus that infects bacteria), Hershey and Chase detected this tracer in the DNA of bacterial cells which the phage had infected. In contrast, the radioactive sulfur atoms they had introduced into the phage’s proteins were not transferred. This suggested that the genetic material which allowed the virus to hi-jack a bacterial cell was indeed DNA.
Meanwhile, Linus Pauling was homing in on the target. In December 1952 his son Peter (then a graduate student at Cambridge) showed Watson a letter indicating that the publication of a new DNA model was imminent. The race seemed over – but when Crick and Watson saw a draft version of Pauling’s paper in January 1953 they detected an obvious flaw in his structure. Realising that others would soon spot it, they intensified their own efforts.
Pauling’s model was another three-chain helix with the phosphates inside. But to permit cross-linking of the chains by hydrogen bonds, Pauling had left the phosphate groups un-ionised. Since the B-form of DNA was heavily hydrated, this seemed as implausible as the salt bridges in the first Cambridge model. The logic underlying both structures was similar, however. The sugar–phosphate backbone was put inside because packing differently shaped base molecules neatly into the centre of the spiral would be very difficult.
Watson quickly brought the news of Pauling’s blunder to King’s. It reinforced Franklin’s conviction that model-building was premature, and when Watson suggested they might collaborate to forestall Pauling she grew distinctly hostile. But Wilkins was more cooperative. He showed Watson one of Franklin’s unpublished X-ray pictures of DNA’s B-form, which indicated that the molecule was helical, although its structural details remained unclear. Watson then began considering the possibility of a two-strand helix with an external sugar-phosphate backbone, and the bases paired like-with-like on the inside of the spiral.Chargaff was amused by Crick and Watson's patchy knowledge of chemistry
Unfortunately, he was unable to manoeuvre the base pairs into positions where hydrogen bonds would form easily between them. Furthermore, since the double-ring molecules of adenine and guanine are larger than the single rings of thymine and cytosine, in a like-with-like structure the two chains had to be set further apart to accommodate the bigger base-pairs, but closer together for the smaller ones. Crick quickly showed that this kinked structure would not fit the crystallographic data. There were also chemical objections to it.
Chemical clues
Crick and Watson were sharing an office with Jerry Donohue, an American chemist on a six-month visit to Cambridge. On seeing Watson’s model he commented that nucleobases can exist in different tautomeric forms. Donohue added that, although standard textbooks depicted thymine and guanine in the enol-form, many chemists thought their keto-forms predominated. This change made hydrogen bonding between similar bases even more problematic.
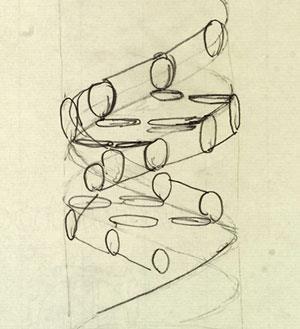
At first, Crick and Watson saw no particular significance in Chargaff’s figures. But in February 1953 Watson suddenly realised that they were compatible with a structure composed of two complementary DNA spirals, in which every adenine molecule on one chain was paired with a thymine molecule on the other (and likewise every guanine molecule with a cytosine one). This greatly improved the fit of the bases, permitting hydrogen bonds to cross-link the two spiral chains. Crick’s crystallographic calculations confirmed that the structure would produce X-ray diffraction patterns like those in Franklin’s pictures.
Having built a three-dimensional model of their structure, Crick and Watson spent several days checking that the lengths and angles of all its chemical bonds were correct, before unveiling it on 28 February 1953. This time, Wilkins and Franklin were convinced. In April, a short paper explaining the double helical structure appeared in Nature, alongside separate but supportive contributions from the King’s group. It included a modest, but momentous, proposition: ‘…the specific pairing we have postulated immediately suggests a possible copying mechanism for the genetic material.’ Many problems remained to be solved – but the revolution had begun.
Further reading
F H C Crick, What Mad Pursuit, New York, Basic Books, 1988
R C Olby, The Path to the Double Helix, New York, Dover, (2nd Ed 1994)
A Sayre, Rosalind Franklin and DNA, New York, Norton, 1975
Watson, J, The Double Helix London, Weidenfeld & Nicholson, 1981
Watson, J, Interview with Brenda Maddox, CSHL Archives http://bit.ly/QuZi6P
Linus Pauling and the race for DNA, a collection of resources from Oregon State University
A (free) collection of Nature papers on DNA from 1953, including:
J D Watson and F H C Crick, Nature, 1953, 171, 737 (DOI: 10.1038/171737a0)
M H F Wilkins, A R Stokes & H R Wilson, Nature, 1953, 171, 738 (DOI: 10.1038/171738a0)
R Franklin and R G Gosling, Nature, 1953 171, 740 (DOI: 10.1038/171740a0)











No comments yet