Fluorescent protein probes can illuminate cells’ inner workings, but making them takes effort – and luck, as Andy Extance discovers
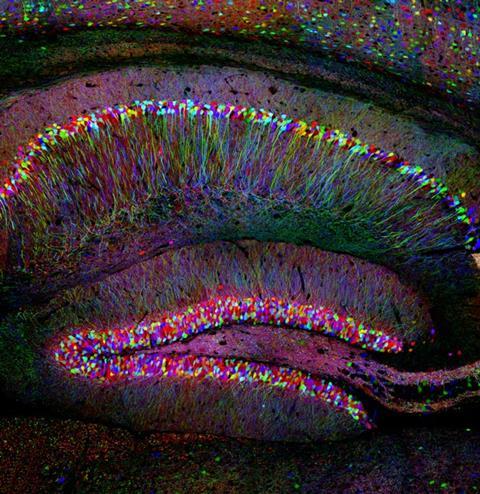
When Joshua Sanes mated a transgenic mouse with red fluorescent neurons to another with green ones, it spawned more than a litter of pups with brains that could glow gaudily. Sanes, a researcher at Harvard University in the US, didn’t just see green and red neurons, some produced both colours and therefore appeared yellow – and that gave postdoctoral fellow Jean Livet an idea. He realised that cells could be made to produce multiple colours in varying amounts, mixing to produce the in-between colours. The ‘stroke of genius’, according to Sanes, came in using the mosaic effect to make it much easier to label and study otherwise hard-to-distinguish cells. Livet and Sanes’ team first published the system that intentionally genetically engineered an optimised version of the effect into mouse neurons in 2007, dubbing the technique ‘Brainbow’.1
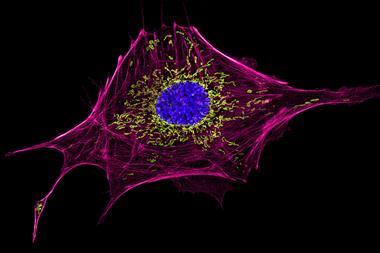
It’s a striking example of how researchers seek to exploit fluorescent protein structures to spotlight exactly what cells do. Since its isolation in 1962, green fluorescent protein (GFP) from the bioluminescent jellyfish Aequorea victoria has illuminated many organisms with its eerie hue. However, despite fluorescent proteins winning the Nobel prize in chemistry in 2008, most sophisticated fluorescent probes seeking to report biological, biochemical and biophysical activity have seen faint success. Sanes admits that even the original version of Brainbow, which is now in its third generation, ‘was maybe less easy to use than we would have liked’. But thanks to continued effort, protein probes today are increasingly setting a shining example, often – as in Livet and Sanes’ case – with a big contribution from chance.
Brainbow inserts genetic instructions for making three or four different colour fluorescent proteins into a specific cell type. Each cell makes randomly varying amounts of each protein, meaning around 100 colours are produced. Brainbow therefore suits long, thin neuron cells well, Sanes explains. ‘If neurons go to different places, you can follow each of them,’ he says, an approach that has helped show how visual systems are wired, for example. Another common use is in watching cell division as animals grow. ‘Say you had 17 cells that are 17 different colours, and wait until you have 1700 cells – they’ll still be only 17 different colours,’ Sanes says. ‘By looking at that you learn a lot about development.’
Many labs now use Brainbow successfully, thanks in part to how his team has resolved a number of colouring puzzles. In particular, the first version of Brainbow ‘was in many cases frankly not bright enough’. ‘Because you’re using different colours every individual colour is a bit less bright,’ Sanes explains. Resolving this issue was possible because GFP’s discovery triggered what the Harvard researcher calls a ‘revolution’ that revealed a wide range of possible fluorescent proteins. While his team could choose the brightest available ‘it would still be nice to have brighter proteins’, he says.
Red alert
Other scientists are trying to help. For many, the prime target is the three out of 238 amino acids in GFP that spontaneously react to form a two-ring chromophore in the presence of oxygen. The rest of the protein forms a barrel-like shape around a single protein thread carrying the three critical amino acids, an arrangement that favours fluorescent chromophore formation. In the two decades since that structure was deciphered, researchers have made small mutations in the genetic code that tells organisms to produce the protein. As well as the results of such efforts to change GFP’s spectral properties, Sanes emphasises that his team also benefited from groups searching for proteins in other fluorescent creatures.
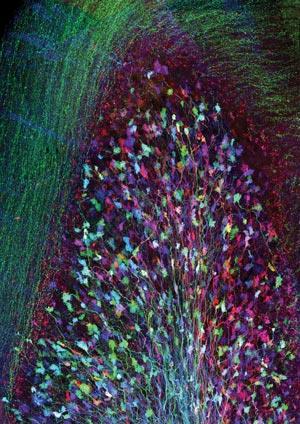
Red-emitting fluorescent proteins have long been a highly prized target for such protein engineers and hunters. Under a microscope, living tissue absorbs and scatters red light less than other colours, and red light harms and excites unwanted fluorescence in the tissue less. However, compared to the green offered by today’s best enhanced GFP, existing deep red proteins are typically dim and more prone to being broken down by light. And while even proteins from other organisms have the same general barrel shape as GFP, their chromophore rings often form less readily.
Ulrich Nienhaus from the Karlsruhe Institute of Technology in Germany is among those who are therefore pursuing new, improved alternatives. Together with Jörg Wiedenmann from the University of Southampton in the UK, Nienhaus discovered that in the bubble-tip anemone Entacmaea quadricolor four molecules of the same protein came together to produce red fluorescence. However, such tetramer-forming sequences can cause unwanted aggregation when added onto other proteins, harming the function scientists want to study.
To produce a more useful protein, Nienhaus, Wiedenmann and their colleagues went through a long series of mutations. First they produced a bright red-emitting non-aggregating monomeric protein, mRuby. From that they developed mGarnet, whose brightness is similar to other red fluorescent proteins, but still not as good as GFP.2 It is very stable when exposed to light though, which is especially important for another Nobel-winning technology, this time 2014’s stimulated emission depletion (STED) super-resolution microscopy. To provide images on the smallest scales, STED uses a focused laser beam that can excite fluorescent proteins, and a second doughnut-shaped laser beam quenches fluorescence at the first spot’s edges. Scanning the beams across a sample builds up a high-resolution picture. This means that proteins get excited and quenched many times, and in turn that their resilience is vital.
The difference in amino acid sequence between mGarnet and mRuby is just four residues, yet the researchers had to produce over 300 different mutants to discover it. ‘We do not have much control yet,’ Nienhaus admits. ‘It is trial and error. We are trying to modify things that have been shown to be successful for other fluorescent proteins, but that may fail completely. We still do not understand the interplay between protein scaffold and chromophore well enough to be able to predict the outcome of such rational engineering approaches.’
Enter the volt
Such a prominent role for fortune is common among advances in protein probes that scientists use to study cells. Another example is protein sensors that detect voltage within a cell, reporting their readings through the intensity of their fluorescence. Until 2012, the best such system offered fluorescence changes that were too small and with a time of response too slow to be easily exploited.
Biosensors for voltage are just now starting to be good enough to be useful
Lawrence Cohen
Nevertheless, Lei Jin of Yale University in the US sought to engineer it into a cell line she could grow reliably in her lab to study the effect of using different fluorescent proteins. Not only did she succeed in this, but the voltage-driven fluorescence change the cells produced was at least five times greater than had ever been seen before.3 The improvement arose thanks to ‘a lucky spontaneous mutation’ in the cell line, explains Lawrence Cohen, who leads the group Jin was working in. He hopes the new sensor, dubbed ArcLight, will help scientists measure the electrical potentials at the core of some vital biological processes without needing electrodes.
‘You can see the potentials with a camera, in many cells at once, which is really hard to do with an electrode,’ Cohen says. ‘You can do much better measurements of activity in hearts and brains. The special thing about protein voltage sensors is that you can make them appear in specific cell types. For example, in the retina, there are about 100 different cell types. If you want to find out how individual cell types participate in generating an output then you need a protein sensor you can get to appear in each cell type, one at a time, as opposed to an organic dye where every cell is activated and binds it.’
Thanks to advances such as ArcLight, ‘biosensors for voltage are just now starting to be good enough to be useful’ for applications like this, according to Cohen. But he adds that ‘until very recently, no-one had any idea’ how voltage influenced proteins’ fluorescence – and therefore further improvements currently pose a significant challenge.
Ion out flaws
Yet such improvements are exactly what Amy Palmer from the University of Colorado in the US wants to see. ‘There are still a large number of sensors that are not widely used, perhaps because they aren’t robust enough for non-experts,’ she explains.
A few notable success stories have already arisen from optimisation efforts, Palmer stresses. She considers the most successful to be single fluorescent-protein-based calcium sensors known as GCaMP, optimised by researchers at Howard Hughes Medical Institute’s Janelia Farm campus in Virginia. Originally developed using components including GFP and the calcium-binding protein calmodulin in 2001, GCaMP is now in its sixth generation.4 The combined protein is dark when calcium is absent, and fluoresces when it’s present.
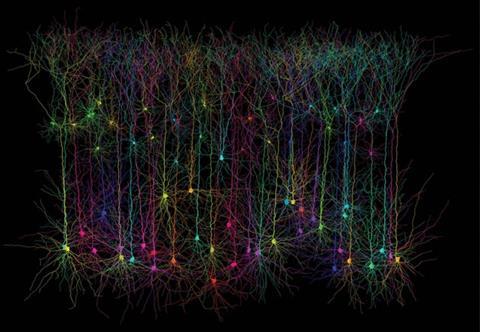
‘These sensors are now incredibly robust and work for a large number of researchers,’ Palmer says. GCaMP is especially useful in monitoring events like molecules binding to proteins at cell surfaces, triggering temporary ‘transient’ changes in calcium ion concentrations inside the cell, she adds. ‘These improvements were the result of effort to optimise the signal, improve stability, and ensure the sensors would function in various model organisms.’
GCaMP is a measure against which Palmer’s team tests the protein calcium sensors it’s developing. Two fluorescent proteins can directly interact through fluorescence resonance energy transfer (Fret) when they get close together and the emission spectrum of one matches the other’s excitation spectrum. With the addition of an ion-sensing section of protein, such pairs can report concentration changes. As calcium ions bind, the Fret sensor protein’s conformation changes, increasing energy transfer, and making the protein receiving the energy fluoresce brightly.
‘Fret sensors can be used to quantify molecules and ions in cells,’ Palmer explains. As well as calcium, her team is also working on monitors for zinc ions, using cyan and yellow fluorescent proteins.5 ‘We and others have developed a toolbox of multiple sensors,’ she adds. ‘These have led to robust estimates of zinc in cell cytoplasm and new hypotheses about how cells use and regulate zinc.’
This could help expand on findings that metal ions in cells change in defined and measurable ways in response to different stimuli, Palmer says. ‘For example, when an oocyte – an egg cell – gets fertilised by a sperm, calcium signals are known to occur,’ she says. ‘With each calcium signal zinc is released from the oocyte. It’s not clear what that zinc is doing yet, but it’s quite striking, so the community is now focused on how to decode these dynamics.’
Leave it to chance
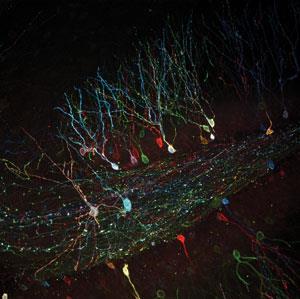
Palmer’s work relies on high-throughput approaches both to comprehensively image the full range of cell responses and to design new sensor systems. In the future, she’s keen to use this method to improve fluorescent proteins’ biophysical properties, including switching between states. ‘If we can figure out what controls switching timing or magnitude, we might be able to optimise them for different applications,’ she says.
The archetype of a fluorescent protein that switches between colours arose from another incidence of serendipity, according to Yoshikazu Imanishi from Case Western Reserve University in Ohio, US. He explains how Atsushi Miyawaki’s team at Riken in Saitama, Japan, was trying to characterise a new green fluorescent protein from a stony coral, Trachyphyllia geoffroyi.6 ‘They left this green fluorescent protein on the bench and the next day they found it had gone from green to red,’ Imanishi says. ‘They decided to call it kaede, which means maple leaf in Japanese, as maple leaves also turn from green to red.’
Kaede’s colour change is caused by ultraviolet light, and therefore it’s referred to as a photoconvertible protein, many more of which have since been developed. Imanishi’s team genetically engineers animals to add such probe groups to particular proteins, and track their behaviour in specific cells. One possibility is looking at what happens to proteins after they’re made. ‘We can convert all the proteins in the cell to red, and after that all the newly produced proteins are green, so we can easily visualise them,’ he says.
Microscopes are 300 years old, yet there’s been a huge amount of development in the last six years
Lawrence Cohen
The Case Western scientists did this to watch proteins move, and test whether actin was involved in carrying key proteins around the cell.7 ‘Actin is a highway for a motor protein that transports small vesicles containing membrane proteins,’ Imanishi says. ‘We destroyed actin filaments chemically, and monitored the transport of the cyclic GMP-gated channel and G-protein coupled receptor. We found that the G-protein coupled receptor can go to the special membrane compartment where it’s supposed to be, but cyclic GMP-gated channels couldn’t go to the plasma membrane. One transport mechanism is dependent on actin but the other isn’t.’
Switchable fluorescent proteins enable another 2014 chemistry Nobel-winning super-resolution microscopy method, photoactivated localisation microscopy (Palm). The method can use green fluorescent proteins that can be flipped from off to on, for example, or irreversibly activated. ‘Visualising single fluorescent proteins requires a very small amount of that protein, because if two fluorescent proteins come together they are difficult to resolve,’ Imanishi says. ‘The idea is to activate fluorescent proteins in very small quantities in a repeated fashion to reconstruct the distributions of many single molecules at resolution below the diffraction limit.’
The involvement in modern microscopy techniques is fitting, given that Cohen cites the history that led up to super-resolution to predict what protein sensors’ future might be. ‘Microscopes are 300 years old, yet there’s been a huge amount of development in the last six years,’ he says. ‘If you devote effort to things they can be made better – that’s a general thing. People will devote effort to making protein sensors, and over a long period of time they’ll get better. I expect protein sensor development’s going to continue for centuries.’
References
1 J Livet et al, Nature, 2007, 450, 56 (DOI: 10.1038/nature06293)
2 A Hense et al, Sci. Rep., 2015, 5, 18006 (DOI: 10.1038/srep18006)
3 L Jin et al, Neuron, 2012, 75, 779 (DOI: 10.1016/j.neuron.2012.06.040)
4 T-W Chen et al, Nature, 2013, 499, 295 (DOI: 10.1038/nature12354)
5 H Ma et al, J. Am. Chem. Soc., 2012, 134, 2488 (DOI: 10.1021/ja2101592)
6 R Ando et al, Proc. Natl Acad. Sci. USA, 2002, 99, 12651 (DOI: 10.1073/pnas.202320599)
7 I Nemet et al, J. Neurosci., 2014, 34, 8164 (DOI: 10.1523/neurosci.1282-14.2014)









No comments yet